Article Text

Abstract
Objective miR-21 was found to be overexpressed in the colon tissues and serum of patients with UC and colorectal cancer (CRC); however, the exact roles of miR-21 in colitis-associated CRC remain unclear. The aim of our study was to investigate the biological mechanisms of miR-21 in colitis-associated colon cancer (CAC).
Design miR-21 expression was examined in the tumours of 62 patients with CRC from China and 37 colitis-associated neoplastic tissues from Japan and Austria. The biological functions of miR-21 were studied using a series of in vitro, in vivo and clinical approaches.
Results miR-21 levels were markedly upregulated in the tumours of 62 patients with CRC, 22 patients with CAC, and in a mouse model of CAC. Following azoxymethane and dextran sulfate sodium intervention, miR-21-knockout mice showed reduced expression of proinflammatory and procarcinogenic cytokines (interleukin (IL) 6, IL-23, IL-17A and IL-21) and a decrease in the size and number of tumours compared with the control mouse group. The absence of miR-21 resulted in the reduced expression of Ki67 and the attenuated proliferation of tumour cells with a simultaneous increase in E-cadherin and decrease in β-catenin and SOX9 in the tumours of CAC mice. Furthermore, the absence of miR-21 increased the expression of its target gene PDCD4 and subsequently modulated nuclear factor (NF)-κB activation. Meanwhile, miR-21 loss reduced STAT3 and Bcl-2 activation, causing an increase in the apoptosis of tumour cells in CAC mice.
Conclusions These observations provide novel evidence for miR-21 blockade to be a key strategy in reducing CAC.
- CANCER
- COLORECTAL CANCER
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Chronic inflammation is an important driver in colorectal cancer (CRC) tumorigenesis and development but the factors related to this neoplastic transformation are poorly understood.
The exact roles of miR-21 in colitis-associated CRC remain unclear.
What are the new findings?
miR-21-KO mice exhibited a reduced production of proinflammatory and procarcinogenic cytokines and decreased tumour size and number compared with control mice.
The absence of miR-21 resulted in attenuated proliferation of tumour cells with a simultaneous increase in E-cadherin and decreased β-catenin and stem cell markers in tumour tissues from colitis-associated colon cancer (CAC) mice.
The absence of miR-21 increased PDCD4 expression, reduced nuclear factor (NF)-κB, STAT3 and Bcl-2 expression in tumour and stromal cells, causing increased apoptosis of tumour cells in CAC.
The current study revealed a crucial involvement of miR-21 in CAC incidence and development.
How might it impact on clinical practice in the foreseeable future?
These observations provide novel evidence for miR-21 blockade to be a key strategy in reducing CAC risk.
Introduction
Chronic inflammation is a driver for the initiation of many diseases,1 ,2 such as UC, a disease with different durations and extent of inflammation that results in colorectal cancer (CRC).2–5 The tumour-promoting cytokines secreted by infiltrated immune cells may contribute to colonic neoplastic transformation and progression.1 ,2 ,6
MicroRNAs (miRNAs) are short non-coding RNAs that participate in diverse biological processes in many species. Emerging studies have demonstrated miRNA-mediated pathway crosstalk between IBD and CRC.7 Cell-type-specific miRNA dysregulation has been reported to be involved in the transition from IBD to CRC.8 ,9 miR-21 has been reported to be differentially expressed in many cancerous diseases. For example, miR-21 is overexpressed in malignant tumour tissues (TTs) of the brain, breast, lung, pancreas, stomach and prostate.10 ,11 Likewise, a higher expression of miR-21 was found in colon adenocarcinomas than in the normal mucosa.12 A number of cancer-related genes, including cell division cycle 25A, phosphatase and tensin homologue (PTEN), tropomyosin 1, programmed cell death protein 4 (PDCD4), protein sprouty homologue 1 and protein sprouty homologue 2 have been reported to be putative targets of miR-21, suggesting that miR-21 functions in regulating cell proliferation and apoptosis.13–17 In addition, miR-21 plays key roles in maintaining T cell effects and regulating chronic inflammatory processes.18 However, the precise mechanisms of miR-21 in promoting the progression of colitis-associated colorectal cancer (CAC) remain to be defined.
Here, we generated miR-21-knockout mice. This model (miR-21 KO) has been clearly described and used in our previous study.19 The mouse model of CAC induced by azoxymethane (AOM, intraperitoneal injection) interspersed with repeated dextran sulfate sodium (DSS, oral administration) has been extensively used. DSS intervention in experimental rodents causes acute inflammation and ulceration in the entire colorectum, which is similar to that observed in human UC.20 In this CAC model, neutralisation of the cytokine tumour necrosis factor (TNF)21 and the transcription factor nuclear factor (NF)-κB reduces tumour proliferation.22 Colon cancer cell growth is induced by T helper type17 lymphocyte- secreted cytokines, such as interleukin (IL) 6, IL-23 and IL-17A.23 ,24 Such cytokines activate signal transducer and activator of transcription 3 (STAT3) in cancer cell, contributing to tumour cell growth, angiogenesis and invasion.25 These findings support the notion that distinct pathways promote CAC, whereas neutralisation of these pathways may help inhibit CAC progression.
Methods
Tissue and serum sample preparation
TTs and adjacent normal tissues from patients with sporadic CRC were collected after curative surgical operations at the Tenth People’s Hospital of Tongji University and the Sixth People’s Hospital of Shanghai Jiao Tong University between 2012 and 2014. The serum samples were collected before the curative surgical operation. We then selected 62 pairs of samples (from 62 patients) that had noticeable inflammation. Among the selected 62 patients, 56 serum samples were collected 1 week before the surgery. In addition, we analysed 53 formalin-fixed, paraffin-embedded (FFPE) colorectal epithelial specimens, which included 37 colitis-associated neoplastic tissues (15 dysplasia cases and 22 cancers) from 37 patients with UC. The UC diagnosis was based on medical history, endoscopic findings, histological examination, laboratory tests and clinical disease presentation. These tissues were retrieved from colectomy specimens that were obtained from Mie University Hospital JAPAN and Medical University of Vienna, Austria. All participants in our study provided written informed consent and willingness to donate their tissue and serum samples for research.
Gene targeting of miR-21 in Mice
miR-21-KO mice (miR-21 flox/flox mice were kindly donated by Prof Zonglai Jiang, School of Life Sciences & Biotechnology, Shanghai Jiao Tong University, China) and wild type (WT) C57BL/6J controls (provided by Jackson Laboratory, Bar Harbor, Maine, USA) were housed in specific pathogen-free conditions with autoclaved food and water. The miR-21-KO mouse construction methods were as follows: the miR-21 flox/flox mice (neo) as described by Zhang et al26 were bred with EIIa-Cre mice (strain: B6.FVB-Tg (EIIa-cre) C5379L mgd/J, The Jackson Laboratory, Bar Harbor, Maine, USA) to generate miR-21 KO mice. Finally, the miR-2 KO mice were interbred to obtain miR-21-KO mice. The genotype of miR-21 flox/flox mice was determined by PCR as described by Zhang et al.26 The genotype of EIIa-Cre mice was determined by PCR as described by Holzenberger et al.27 miR-21-KO mice were detected using the following primers: 5′-CAGAATTGCCCAGGCTTTTA-3′ (mir-21-p1); 5′-AATCCATGAGGCAAGGTGAC-3′ (mir-21-p2).
CAC modelling
CAC modelling was performed as described previously.2 ,28 Briefly, 8- to 10-week-old male WT and miR-21-KO mice were first given an intraperitoneal administration of AOM (A5486, Sigma-Aldrich; 12 mg/kg). On day 7 after AOM intervention, the mice were treated with drinking water including 2% DSS (molecular weight (MW) 36 000–50 000 Da; MP Biomedicals) for the following five consecutive days. On day 20 after the first AOM intervention, the mice received a second intervention of AOM (12 mg/kg). At 7 days after the second AOM intervention, the mice were again given 2% DSS for 5 days, followed by regular water until day 81. During the treatment course, the mice were monitored for body weight, diarrhoea and haematochezia. On day 31 and day 81, colon tissues were opened longitudinally and measured, and the tumours were quantified. Colon sections were frozen in liquid nitrogen and fixed in 10% formalin.
Scoring of disease activity index and histopathology
Scoring of the disease activity index was performed according to previous reports.29 ,30 The mouse disease activity index (ranging from 0 to 4) was calculated as follows: stool consistency; the presence or absence of haematochezia; and weight change. The inflammation was scored on day 31 and day 81. Colon segments were fixed in 10% formalin, paraffin-embedded, cut into 5-µm sections, stained with hematoxylin and eosin (H&E) for microscopic examination and imaged (Nikon Eclipse 80i, Japan). The slides were reviewed and scored for inflammation by two pathologists blinded to the treatment based on the criteria as described:29 score 0, no inflammation; score 1, modest numbers of infiltrating cells in the proper layer; score 2, infiltration of inflammatory cells with mild mucosal hyperplasia; score 3, massive infiltration of immune cells with disrupted mucosal architecture, and marked mucosal hyperplasia; score 4, all of the earlier described plus crypt abscesses or ulceration.
Immunohistochemistry
FFPE specimens (5-µm thick) were routinely deparaffinised in xylene and an ethanol gradient, incubated in 3% hydrogen peroxide for 10 min to block endogenous peroxidase, treated with 10 mM Na citrate buffer, pH 6.0, in a pressure cooker at 120°C for 10 min and then cooled to room temperature. Sections were incubated with the following antibodies: anti-CD3 (rabbit polyclonal, 1:300 dilution; Epitomics); anti-CD68 (rabbit polyclonal, 1:100 dilution; Abcam); anti-Ki67 (rabbit polyclonal, 1:200 dilution; Abcam); β-catenin (rabbit polyclonal, 1:500 dilution Epitomics); anti-SRYbox-containing gene 9 (SOX-9; rabbit polyclonal, 1:100 dilution; Abcam); anti-STAT3 (rabbit polyclonal, 1:100 dilution; Santa Cruz); anti-p-STAT3(Tyr705; rabbit polyclonal, 1:400 dilution; Cell Signaling Technology); anti-Bcl-2 (rabbit polyclonal, 1:200 dilution; Abcam); anti-E-cadherin (rabbit polyclonal, 1:200 dilution; Cell Signaling Technology); and anti-PDCD4 (rabbit polyclonal, 1:400 dilution; Abcam). The primary antibodies were detected using appropriate secondary antibodies (1:500 dilutions, Abcam) at 37°C for 30 min. The 5-µm sections were counterstained with haematoxylin (Abcam), dehydrated and cover-slipped.
Proliferation and apoptosis
The 5-µm FFPE colon sections were assessed through immunohistochemical staining using an anti-Ki67 (rabbit monoclonal, 1:300 dilution; Abcam) antibody. Apoptosis was detected using the In Situ Cell Death Detetion Kit (Roche 11684817910) according to the manufacturer's instructions. Anti-Ki67-positive cells or terminal deoxynucleotidyl transferase (TdT)-mediated nick end labeling (TUNEL)-positive cells were counted by two pathologists blinded to the treatment on 10 random fields at 400× magnification in each slide.
Enzyme-linked immunosorbent assay
The protein levels of TNF-α, IL-6 and IL-17A were measured in the sera of patients with CRC using Sandwich ELISA kits (Anogen, Canada) according to the manufacturer's instructions. The mouse serum protein levels of TNF-α, IL-6, IL-17A and IL-21 were measured at day 31 and day 81 using Sandwich ELISA kits (R&D, USA).
RNA extraction and quantitative real-time PCR
Total RNA from frozen tissues was extracted using Total RNA Extraction Kit (SLNco, Cinoasia, China), and the Reverse Transcription Kit (TOYOBO, Japan) was used for the first strand synthesis of cDNA. Human and murine primers are described in online supplementary table S1. The mRNA levels of TNF-α, IL-6, IL-21 (using glyceraldehyde 3 phosphate dehydrogenase (GAPDH) as endogenous control for normalisation) and miR-21 expression (using U6 as endogenous control for normalisation) in human samples and TNF-α, IL-6, Cox-2, keratinocyte chemoattractant RNA and miR-21 expression in murine colon tissues were assessed using a TaqMan assay (Applied Biosystems, Foster City, USA).
Western blotting
Total proteins of frozen tissues of mice were extracted using lysis buffer (Mammalian Protein Extraction Reagent (M-PER); The following four words (Mammalian Protein Extraction Reagent) is the expansion for the M-PER, Pierce),31 separated on an sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel, and incubated (1:1000 dilution) with anti-E-cadherin, anti-β-catenin, anti-SOX-9, anti-STAT3, anti-p-STAT3, antiphospho-NF-κB p105 (Ser933; rabbit monoclonal; Cell Signaling Technology), antiphospho-NF-κB p65 (Ser468; rabbit polyclonal; Cell Signaling Technology), anti-Bcl-2 and anti-PDCD4 antibodies. An anti-GAPDH antibody (1:8000 dilution; Abcam) was used as an internal control.
Bioinformatics analysis of potential miR-21 targets
The miRecords resource (http://mirecords.umn.edu/miRecords/)32 was used to search for the potential targets of miR-21.
Luciferase reporter assay
Oligonucleotides containing the WT or mutant (MT) target site of the 3′-untranslated regions (3′-UTR) of the PDCD4 were ligated into the pGL3-basic vector (Promega, Wisconsin, USA). Product identification was performed using restriction enzyme digestion (Kpn I and Hind III; Promega) and sequencing. The primers used to amplify the PDCD4 3′-UTR were as follows: WT-forward, 5′-GGGGTACCAACCTCTTCTTAAGTGGAATATTC-3′, WT-reverse, 5′-CCCAAGCTTACTCCCAAAGCACAGTATCTCAA-3′, MT-forward, 5′-GGGGTACCAACCTCTTCTTAAGTGGAATATTCTACAGTTAAACCTT-3′ and MT-reverse, 5′-CCCAAGCTTACTCCCAAAGCACAGTATCTCAA-3′. The LoVo cells were cotransfected with the PDCD4 vector or control, miR-21-mimics, miR-21-inhibitor or a negative control with Lipofectamine 2000 (Invitrogen). After 36 h, luciferase assays were performed normalising to the Renilla luciferase using the Dual-Luciferase Reporter Assay System (Promega).
Statistical analyses
The results are presented as the mean±SEM. Multiple correlation analysis was assessed by Pearson's test using GraphPad Prism software (V.5.00, San Diego, California, USA). Significant differences between categorised groups were analysed by Student's t test or Kruskal-Wallis tests, as appropriate. The χ2 test and Fisher's exact test were used where appropriate to analyse the relationship between miR-21 levels and the clinicopathological parameters using SPSS software (V.15.0, Chicago, Illinois, USA). A p value <0.05 (two-sided) was considered statistically significant.
Results
miR-21 overexpression in human CRC and mouse CAC
miR-21 expression was more pronounced in the cancer tissues of 62 patients with sporadic CRC than in non-tumour mucosae (normal control; figure 1A). There was an obvious correlation between miR-21 levels and depth of invasion in patients with CRC (p=0.038) but no significant correlation between miR-21 levels and sex, age, tumour localisation, tumour size, tumor-node-metastasis (TNM) staging, lymph node invasion, distant metastasis or differentiation in patients with CRC (table 1). Inflammatory infiltration was present in the non-tumour mucosa and TT of patients with CRC (figure 1B); therefore, we analysed the mRNA levels of TNF-α, IL-6 and IL-21 in CRC samples (n=62). miR-21 expression significantly correlated with the mRNA levels of TNF-α, IL-6 and IL-21 in CRC samples (figure 1C–E). Accordingly, the protein levels of TNF-α (n=56), IL-6 (n=56) and IL-17A (n=56) in the serum samples of patients with CRC significantly correlated with miR-21 expression (figure 1F–H). Furthermore, to determine whether miR-21 dysregulation is consistent with colitis-associated cancer, we quantified the expression analysis using FFPE specimens from patients with UC-associated dysplasia (n=15) and cancer (n=22). As expected, miR-21 expression showed a stepwise increase in dysplasia samples (p<0.05) and cancer (p<0.05) compared with normal colonic epithelium (figure 1I). Collectively, these results suggested that miR-21 might be intimately involved in inflammation-induced CRC. Based upon these findings, we focused on the assessment of the biological impact of miR-21 in colitis-associated carcinogenesis.
Association of miR-21 expression with the clinicopathological features of patients with colorectal cancer
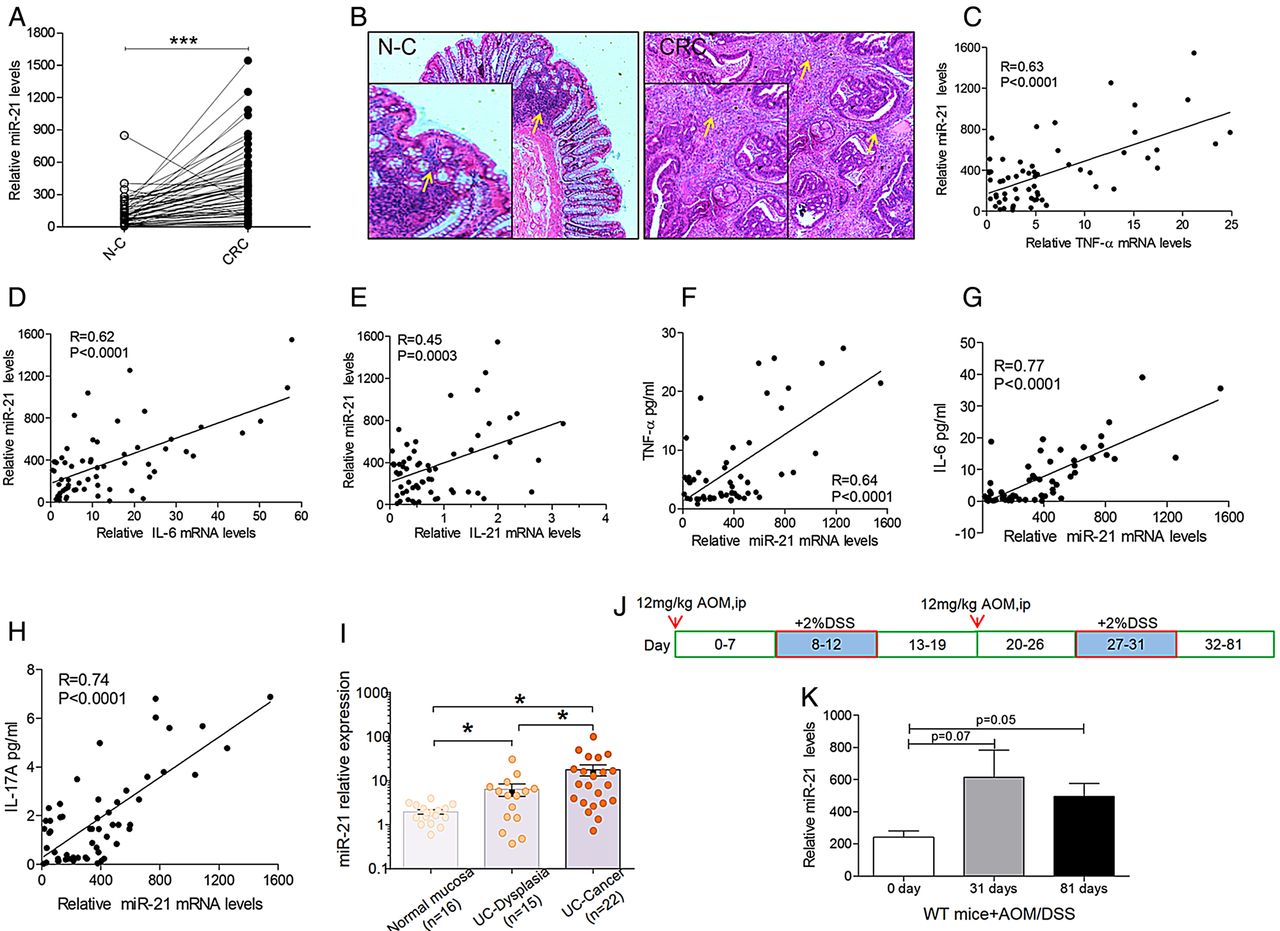
miR-21 is overexpressed in patients with colorectal cancer (CRC) and mice. (A) miR-21 in fresh colonic specimens taken from adjacent tumoral (CRC) and non-tumoral (normal control (N-C)) areas of 62 patients with sporadic CRC were quantified by qPCR (mean±SEM, n=62, ***p<0.001). (B) Inflammatory infiltration is present in the colon of patients with sporadic CRC by H&E stain (original magnification 400×, the yellow arrows indicate the inflammatory infiltration of the colon). There was a significant correlation between miR-21 expression and mRNA levels of (C) tumour necrosis factor α (TNF-α), (D) interleukin (IL)-6 and (E) IL-21 in CRC samples (n=62). Accordingly, miR-21 expression significantly correlated with serum protein levels of (F) TNF-α (n=56), (G) IL-6 (n=56) and (H) IL-17A (n=56; p<0.0001). The inset shows the Pearson's r correlation and corresponding p value. (I) Expression levels of miR-21 in normal mucosa (n=16), UC-associated dysplasia (n=15) and UC-associated cancer (n=22). The Y-axis represents the relative expression of miRNAs normalised to RNU-6 expression. Significant differences were determined using Kruskal-Wallis tests (mean±SEM, *p<0.05). (J) Diagram of azoxymethane/dextran sulfate sodium (AOM/DSS) administration during the experiment to establish the colitis-associated colon cancer model. (K) miR-21 overexpression in the colon of wild type (WT) mice on day 31 and day 81 of AOM/DSS administration compared with on day 0 (mean±SEM, n≥4, *p<0.05).
We next detected the levels of miR-21 in AOM/DSS-induced CAC. WT mice were administered two cycles of DSS/AOM treatment for these experiments (figure 1J). We sacrificed WT and KO mice on day 31 and day 81, after which, the colon was resected, and the expression level of miR-21 was detected. After AOM/DSS administration, miR-21 expression in the colon was significantly increased compared with miR-21 levels before AOM/DSS administration in WT mice (figure 1K). Therefore, the WT mice that were treated with AOM/DSS had significantly increased expression of miR-21.
miR-21 KO mice were resistant to CAC
To decipher miR-21 function in CAC, we used miR-21-KO mice, in which the miR-21 gene was knocked out systemically. The mice appeared fertile and as healthy as WT mice in standard feeding conditions. When the animals in this model were exposed to the AOM/DSS treatment, the mice exhibited less weight loss, especially after DSS treatment, than the WT mice (figure 2A, C). Associated with the observed weight loss ensuing from the DSS treatment, the miR-21-KO mice also showed a decreased disease index as evaluated by the detection of rectal bleeding and diarrhoea scores (figure 2B, C). At the end of the protocol, miR-21-KO and WT mice were euthanised, and the colons were resected to measure the tumour number and colon length. The tumour incidence was 100% in the miR-21-KO and the WT mice, but the miR-21 KO mice displayed increased colon length (figure 2D, E) and a noticeable decrease in the number of macroscopically visible tumours compared with WT mice (figure 2E–G). Furthermore, the tumours from the miR-21-KO mice were markedly smaller in size, with approximately 50% of miR-21-KO mice developing adenomas or tumours smaller than 1 mm in diameter (figure 2H).

miR-21-KO mice display decreased tumorigenesis and growth upon azoxymethane/dextran sulfate sodium (AOM/DSS) administration. (A) Body weight and (B) disease index were determined during the experimental procedure (n≥10) and (C) showed significant increases in body weight and attenuated rectal prolapse in miR-21-KO mice. (D) Colon length of wild type (WT) and miR-21-KO mice were examined (mean±SEM, n=10, **p<0.01) and (E) exhibited shortening after 81 days of AOM/DSS exposure. (F) Tumour numbers were counted at the end of the protocol (mean±SEM, n=10, *p<0.05), and (G) significantly fewer tumours were found in miR-21-KO mice than in WT mice. (H) Tumour sizes were measured, and a histogram shows the tumour size distribution (n=10).
miR-21 KO attenuates colitis
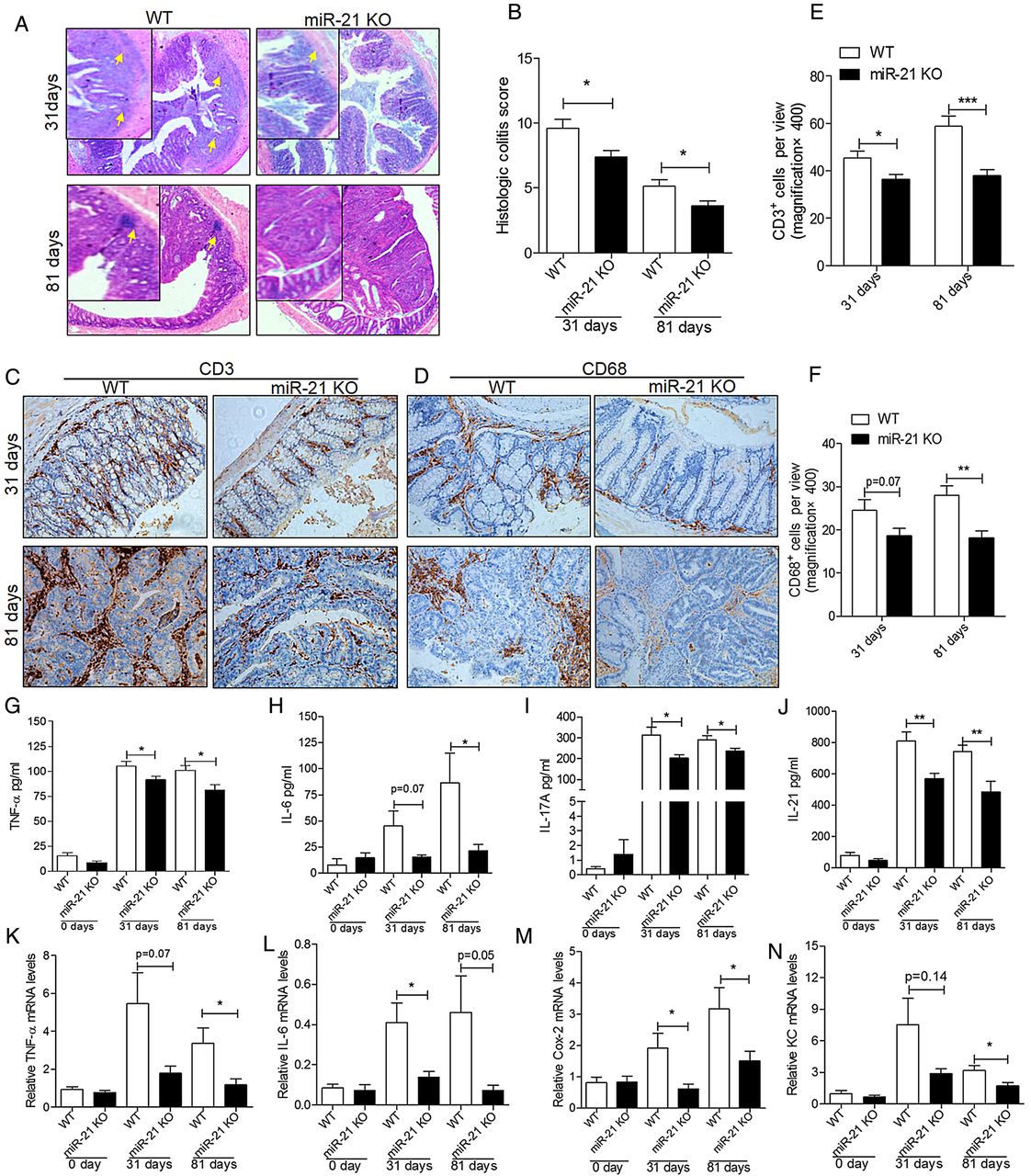
To further investigate the influence of colitis on CAC, we evaluated the colitis and the injury severity scores. Histological analysis of colon sections on day 31 and day 81 after AOM/DSS treatment showed reduced inflammatory and tissue injury indices in knockout mice than in WT mice (figure 3A). Moreover, histological scores of colitis showed marked differences in inflammation scores (figure 3B). Furthermore, to detect cell-specific differences in the following inflammatory response, immunohistochemistry (IHC) analyses were performed to detect CD68+ and CD3+ cells in the colon on day 31 and day 81. The IHC staining of CD3 (figure 3C) and CD68 (figure 3D) showed that CD3+ and CD68+ cells were significantly reduced in the colon of miR-21-KO mice than in WT mice (figure 3E, F). These data showed that the tissue inflammation induced by AOM/DSS was reduced under miR-21 deficiency.
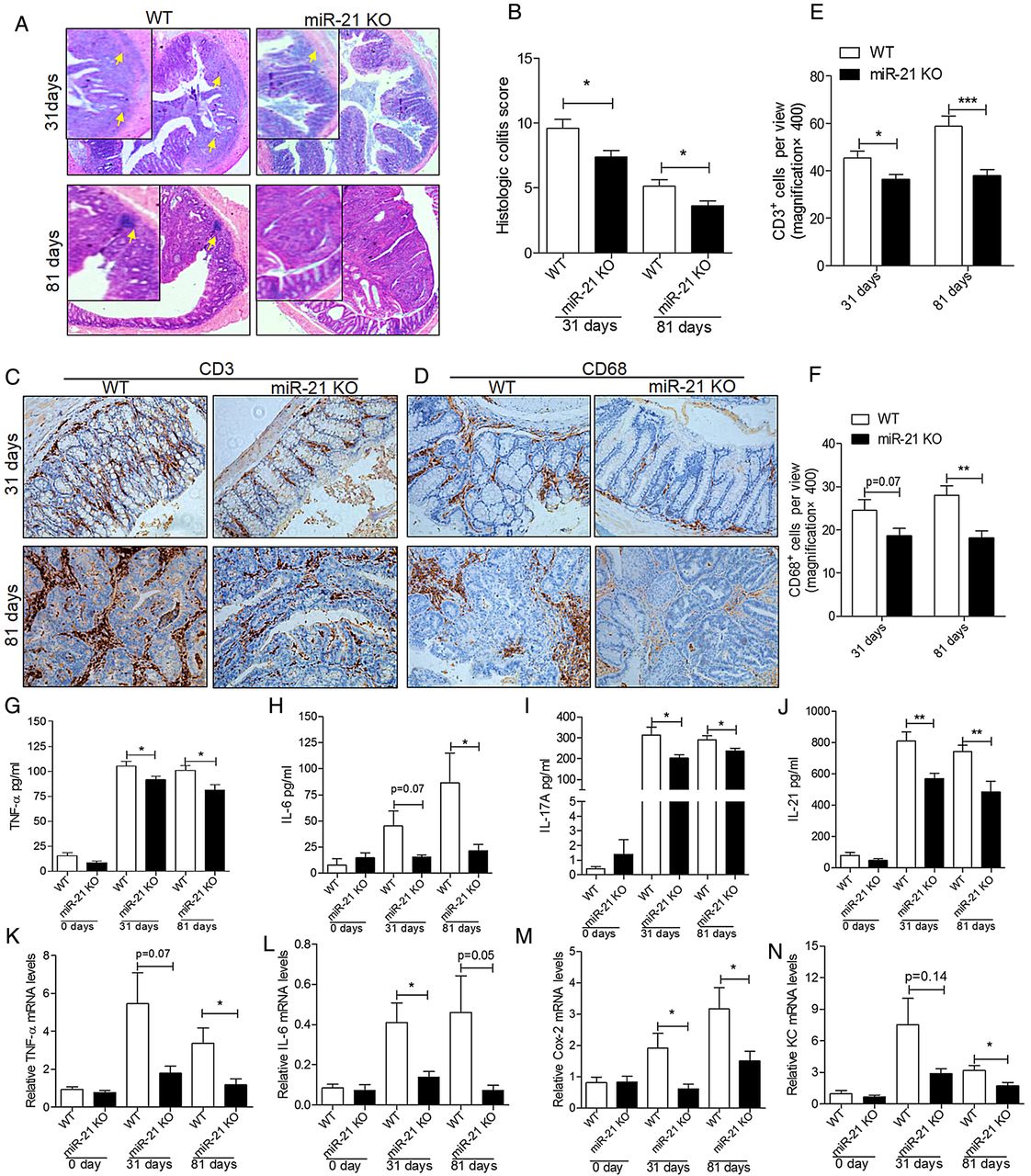
miR-21-KO mice show milder colitis and produce less proinflammatory and procarcinogenic cytokines than wild type (WT) mice after azoxymethane/dextran sulfate sodium (AOM/DSS) treatment. (A) Representative H&E images of colon colitis of miR-21-KO and WT mice on day 31 and day 81 of AOM/DSS treatment (original magnification 400×, the yellow arrows indicate the inflammatory infiltration and the injured colon). (B) Histological colitis score displays decreased colitis and injury of miR-21-KO mice (mean±SEM, n≥5, *p<0.05). Representative IHC images of (C) CD3 and (D) CD68 in miR-21-KO and WT mice on day 31 and day 81 of AOM/DSS treatment (original magnification 200×). Quantification of (E) CD3-positive (CD3+) and (F) CD68-positive (CD68+) cells per view (400×, n=10) showed that CD3+ and CD68+ cells were significantly decreased in the colon of miR-21-KO mice (mean±SEM, ***p<0.001). The serum levels of (G) tumour necrosis factor α (TNF-α), (H) interleukin (IL)-6, (I) IL-17A and (J) IL-21 were increased in WT mice more than in miR-21-KO mice at 31 days and 81 days of AOM/DSS treatment (mean±SEM, n≥5; *p<0.05, **p<0.01, respectively). Meanwhile, mRNA expression levels of (K) TNF-α, (L) IL-6, (M) Cox-2 and (N) keratinocyte chemoattractant (KC) were also significantly increased in the colon of WT mice compared with miR-21-KO mice at 31 days and 81 days of AOM/DSS treatment (mean±SEM, n≥4, *p<0.05, Student's t test).
miR-21-KO mice produce less proinflammatory and procarcinogenic cytokines
The contribution of immune cells to CAC development may be through cytokine production;25 therefore, we examined the cytokine profile of the model mice. Analyses of cytokine proteins, including TNF-α, IL-6, IL-17A and IL-21, by ELISA confirmed that these cytokines were increased in WT mice compared with miR-21-KO mice after 31days and 81 days of AOM/DSS treatment (figure 3G–J). Accordingly, there was a significant increase in TNF-α, IL-6, Cox-2 and keratinocyte chemoattractant transcripts in the colon of WT mice compared with miR-21-KO mice after AOM/DSS treatment (figure 3K–N). Because IL-17A is mainly produced by T cells, whereas IL-6 is mostly produced by T cells and macrophages,2 these findings indicate that the reduced tumorigenesis of knockout mice is related to the reduced production of IL-6,TNF-α, IL-17A and IL-21 by a diminished infiltration of T cells and myeloid cells into the tumour.
miR-21 modulates CAC cell proliferation and enhances the expression of CAC surrogate markers
To further distinguish the effects of the deregulated carcinogenic signals in AOM/DSS-treated mice, we analysed basic parameters that correlate to the tumorigenic process in the colon. The proliferation of miR-21-KO mouse colon cells was significantly decreased before and after AOM/DSS treatment (0, 31 and 81 days), as assessed by Ki67 staining (figure 4A, B). This finding is in agreement with the reduced tumour development in miR-21-KO mice shown in figure 2. We also found a significantly increased expression of E-cadherin, a CAC surrogate marker (figure 4C, F), and reduced expression of β-catenin (figure 4D, F) and SOX9 (figure 4E, F) in TTs of miR-21-KO mice. Taken together, these findings indicate that miR-21 influences epithelial cell proliferation and modulates the procarcinogenic environment in CAC TTs.

miR-21-KO mice display inhibited proliferation in tumour tissues (TTs) and altered expression of tumour-associated genes. Paraffin-embedded colon sections from wild type (WT) and miR-21-KO mice after azoxymethane/dextran sulfate sodium (AOM/DSS) initiation were stained with Ki-67 for assessment of proliferation. (A) Representative images of Ki-67 stain on day 0, day 31 and day 81 of AOM/DSS treatment (original magnification 200×). (B) Quantification of Ki-67-positive cells per view (400×, n=10) showed Ki-67-positive cells were significantly decreased in the colon of miR-21-KO mice compared with that of WT mice (mean±SEM, ***p<0.001). (C) Representative images from immunostaining of normal tissue (NT) and TT samples (n=6) on day 81 of the experimental protocol with antibodies against E-cadherin (original magnification 200× and 400×). Representative images from IHC of colon tissues (n=5) on day 0, day 32 and day 82 of the protocol with antibodies against (D) β-catenin and (E) SOX-9 (original magnification 200×). From the IHC images, there were significant differences in expression in TT, but no significant differences in the NTs. (F) Western blotting further confirms that E-cadherin expression was significantly increased, and β-catenin and SOX-9 expression was significantly decreased in TT (on day 81) of miR-21-KO mice compared with WT mice. (G) The expression ratio of E-cadherin, β-catenin and SOX-9 proteins over GAPDH in TT of miR-21-KO mice and WT mice (mean±SEM, n=5, *p<0.05, **p<0.01).
miR-21 targets PDCD4 and decreased NF-κB activation
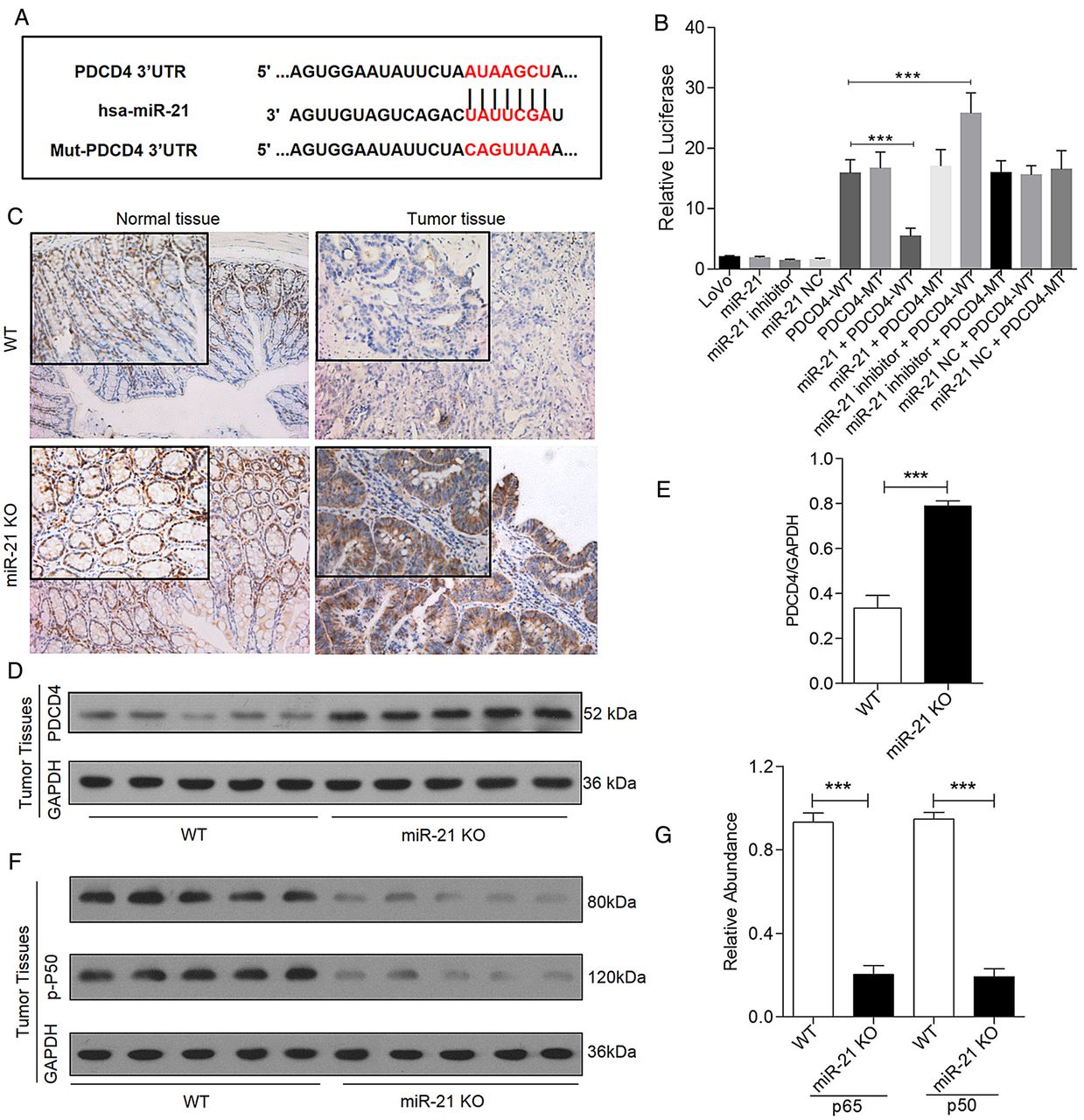
Using the miRecords resource website32 and the limitation of a hit in at least 4 of the 11 bioinformatics prediction tools, eight potential target genes of miR-21 were found to meet this criterion (see online supplementary table S2). We selected PDCD4 for analysis because it showed the highest frequency. In addition, several studies demonstrated that PDCD4 serves as a putative target of miR-21, and PDCD4 inhibits tumour proliferation through modulating NF-κB activity.14 ,33 Therefore, we conducted a luciferase reporter assay combined with western blotting assays to investigate whether miR-21 targets PDCD4 and modulates NF-κB activation. A plasmid harbouring the WT/MT of the predicted region at the 3′UTR of PDCD4 region was constructed (figure 5A). The cotransfection of LoVo cells with the WT reporter plasmid and miR-21 decreased the relative luciferase activity (figure 5B). In addition, decreased PDCD4 protein levels were detected in the normal and tumour colon tissues of WT mice compared with the KO mice (figure 5C–E). Because NF-κB activation in intestinal epithelial cells (IECs) promotes the survival of newly emerging premalignant cells,22 ,23 and the PDCD4/p65 interaction prevents NF-κB-dependent transcription in cancer,33 we further measured NF-κB activation by examining phosphorylated-NF-κB p105 (p-p50) and phosphorylated-NF-κB p65 (p-p65) in colonic TT derived from mice sacrificed on day 81. The western blotting analyses showed significantly decreased expression of p-p50 and p-p65 in the TT of miR-21-KO mice but not control mice (figure 5F, G).

miR-21 targets programmed cell death protein 4 (PDCD4) and decreased nuclear factor (NF)-κB activation in colitis-associated colon cancer (A) miR-21 and the wild type (WT) or mutant (MT) target region in the PDCD4 3′-UTR. The mutated binding site is marked in red. (B) The luciferase activity with or without miR-21 in LoVo cells. ***p<0.05. The miR-21 combined vector containing the WT PDCD4 3′-UTR versus vector combined WT PDCD4 3′-UTR; miR-21-inhibitor combined vector containing the WT PDCD4 3′-UTR versus the vector containing the WT PDCD4 3′-UTR. (C) Representative immunostaining of normal tissues (NTs) and tumour tissues (TTs) on day 81 of the experiment with antibodies against PDCD4 (original magnification 200× and 400×, n=5). (D and E) Western blotting of PDCD4 in colon TTs normalised to GAPDH (mean±SEM, n=5, ***p<0.001). (F and G) Western blotting showing the decrease in p-NF-κB p105 and p-NF-κB p65 expression in TTs of miR-21-KO mice (mean±SEM, n=5, ***p<0.001).
miR-21 KO reduced STAT3 activation and increased tumour cell apoptosis in CAC
The cytokines IL-6, IL-17A and IL-21 are powerful activators of the transcription factor STAT3,2 ,34 ,35 a crucial modulator of inflammation.2 ,36 We thus compared STAT3 and phosphorylated (p)-STAT3 (Tyr705) in colon TT derived from mice sacrificed on day 81. Activation of STAT3 signalling was verified by western blotting, with increased p-STAT3 and nuclear p-STAT3 bands in colonic tumour extracts from WT mice compared with miR-21-KO mice (figure 6A, B). A higher number of STAT3-positive (STAT3+) (figure 6C, D) and p-STAT3+ (figure 6E, F) cells were observed in the TTs of WT mice than in those of miR-21-KO mice, but there was no difference in normal tissues. Cells positive for STAT3 and p-STAT3 were observed in the stroma and epithelium of TTs (figure 6C, E). Consistent with these findings, enhanced expression of the antiapoptotic protein Bcl-2, a target of STAT3,37 was observed in WT mice (figure 6G, H). Accordingly, apoptosis of tumour cells was increased in miR-21-KO mice (figure 6I, J).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Decreased signal transducer and activator of transcription 3 (STAT3) activation and increased apoptotic cells were observed in the tumour tissues (TTs) of miR-21-KO mice. (A) Western blotting showing the decreased expression of STAT3 and p-STAT3 in TTs of miR-21-KO mice. (B) The expression ratio of nuclear p-STAT3 proteins over GAPDH in the TTs of miR-21-KO mice and wild type (WT) mice (mean±SEM, n=5, ***p<0.0001, Student's t test). Representative IHC images of normal tissue (NT) and TT samples (n=6) on day 81 of the experimental protocol with antibodies against (C) STAT3 and (E) p-STAT3 (original magnification 200× and 400×). Quantification of (D) STAT3-positive (p-STAT3+) and (F) p-STAT3-positive cells per view (400×, n=10) showed STAT3+ and p-STAT3+ cells were significantly decreased in colon of miR-21-KO mice compared with control mice (mean±SEM, *** p<0.001). Meanwhile, the TTs of miR-21-KO-mice showed significantly decreased expression of the antiapoptotic protein Bcl-2 and increased apoptosis of tumour cell. (G) Representative images from immunostaining of TT samples (n=6) on day 81 of the experimental protocol with antibodies against Bcl-2 (original magnification 200× and 400×). (H) Western blotting further confirmed the significant decrease of Bcl-2 expression in TTs of miR-21-KO mice. (I) Representative images of TUNEL staining in TTs (original magnification 200× and 400×). (J) The number of TUNEL-positive cells per field (400×, n=10) was significantly higher in colonic TTs of miR-21-KO mice than in WT mice (mean±SEM, ***p<0.001). (K) Model of miR-21 interactions in colitis-associated colon cancer (CAC).
Discussion
The aim of our current study was to clarify the role of miR-21 in the progression of colitis-induced colon tumorigenesis. In a previous study, we showed miR-21 overexpression in the colonic mucosa of patients with UC and IL-10-KO mice19 and found miR-21 upregulation in patients with UC through targeting the Rho GTPase RhoB, which impairs intestinal epithelial barrier function.38 Here, we showed miR-21 overexpression in the neoplastic areas of patients with CRC, and miR-21 expression in neoplastic areas significantly correlated with the expression of TNF-α, IL-6, IL-17A and IL-21. In addition, we analysed the relationship between the clinicopathological characteristics of patients with CRC and miR-21 expression in CRC samples. As expected, miR-21 overexpression correlated with a greater depth of invasion. Furthermore, our data also revealed a stepwise increase in miR-21 expression in UC-associated neoplastic tissues, raising the possibility that miR-21 plays a key role in inflammation-induced colon tumorigenesis. We then extended our research using a well-accepted animal model of CAC.2 ,23 ,28 In the mice, repeated AOM treatment followed by DSS cycles leads to chronic inflammation and the development of colon tumours. After miR-21 knockout, colonic inflammation was decreased, as was the tumour incidence and size.
Our investigation showed that the absence of miR-21 was accompanied by a noticeable reduction in the number of Ki67-positive cells in CAC, whereas the apoptotic cells were significantly increased in CAC TTs. A large number of studies have found that miR-21 potentially contributes to carcinogenesis by compromising DNA damage-induced checkpoint function and cell cycle progression under those conditions, which can lead to chromosomal instability that promotes tumorigenesis.13 ,39 Furthermore, the cancer-stimulating effect of inflammation is now extensively accepted and better understood.40 ,41 Immune cells, which frequently infiltrate tumours and preneoplastic lesions, produce many types of chemokines and cytokines that propagate a localised inflammatory response and promote the growth and survival of premalignant cells by triggering transcription factors such as NF-κB.42 ,43 miR-21 upregulation also activated NF-κB and extracellular-regulated protein kinase signalling in malignant cells suggested a signal amplification effect between miR-21 and cytokines.44
The cytokines or growth factors, such as IL-6, produced upon NF-κB activation in intestinal myeloid cells promote the proliferation and survival of the premalignant IECs generated during early stages of CAC tumorigenesis.23 The cascade of NF-κB-IL-6-STAT3 is a key regulator of the survival and proliferation of tumour-initiating IECs.23 In myeloid cells, inactivation of NF-κB inhibited the production of cytokines such as TNF-α and IL-6 and prevented IEC proliferation during CAC induction.22 The interactions of transcription factors with miRNA are likely to participate in the oncogenic process, and differentially regulated miRNAs are important for this process.7–9 As a transcription factor, STAT3, which is activated by IL-6, directly activates miR-21.45 ,46 miR-21 inhibits tumour suppressors (such as PDCD4 and PTEN),15 resulting in augmented NF-κB activity, which is required to maintain the oncogenic process.45 STAT3 is a target of IL-6, PTEN and miR-21, suggesting a positive feedback loop that relates to the inflammation-cancer switch.45
Here, our data suggest that miR-21 stimulates the development of colon cancer during inflammation. The miR-21-KO colon exhibited markedly reduced activity of NF-κB via the enhanced expression of PDCD4, depressed inflammatory infiltration and produced less tumour-associated inflammatory cytokines, such as TNF-α, IL-6, IL-17A and IL-21.2 ,21 ,36 ,47 The IL-21 produced in excess during colitis promoted the production of IL-17A and IL-6, magnifying colonic inflammation and supporting tumorigenesis.2 Correspondingly, mice lacking IL-6 in bone marrow-derived cells exhibit fewer tumours with a substantial reduction in overall CAC tumour load, whereas the role of immune cell IL-6 appears greater in cancers associated with underlying chronic inflammation.48 ,49 Meanwhile, TNF-α was required to activate a tumour-promoting immune response. The incidence of AOM/DSS-induced CAC was strikingly lower in mice receiving a TNF inhibitor or those lacking the TNF receptor p55.21 Furthermore, the major protumorigenic IL-6 effector is STAT3, whose specific ablation in intestinal epithelial cells interferes with tumour formation and growth and decreased tumour multiplicity in a mouse model of CAC.2 ,23 ,36 Accordingly, we also found that p-STAT3 (Tyr705) was strikingly decreased in the TT of miR-21-KO mice. In addition, the expression of antitumour and tumorigenic proteins and genes, such as E-cadherin, β-catenin and SOX-9, were significantly altered in the TT of AOM/DSS-treated miR-21-KO mice, whereas E-cadherin-based adhesion plays an important role in the development and progression of CRC and modulates the epithelial-mesenchymal transition of CRC cells. Therefore, the altered expression of these genes may be additional factors that restrain CAC development and progression in miR-21-KO mice. However, it is difficult to determine experimentally whether miR-21 is simply increasing inflammation or whether it functions through some other means of increasing neoplasia. In addition, it is a question whether the miR-21 modulation benefit originates from inflammatory cells or epithelial cells. In the future, we hope to further analyse the potential biological function of miR-21 as a therapeutic target for CAC.
In conclusion, this study reveals the crucial involvement of miR-21 in the regulation of the incidence and development of CAC triggered by colitis (figure 6K). miR-21 plays a pathogenic role in chronic inflammatory processes that are associated with augmented CRC risk, providing novel evidence for the miR-21 blockade in reducing the risk of CAC.
Acknowledgments
The authors thank Prof Zonglai Jiang for provision of miR-21 flox/flox mice, and Shanghai Jiao Tong University Affiliated Sixth People's Hospital (Animal Experiment Center, Central Laboratory and Pathology Department) for technical assistance during this study.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Data supplement 1 - Online table S1
- Data supplement 2 - Online table S2
Footnotes
CS and YY contributed equally.
Contributors HQ, YM and AG were the principal investigators and supervised and financed the study. CS, YX, JY, PZ, FW, HH wrote the protocols and (with WW, RG, YO, HC, YL, CG, YY) analysed the data and interpreted the findings. YM, HQ and AG reviewed the study implementation and contributed to the data monitoring. All of the authors had full access to the primary data and the final analysis, and all have seen and approved the final version of the manuscript.
Funding This work was financially sponsored by grants from the National Natural Science Foundation of China (No.81230057; No.81472262; 81200264; No.81372615), the National High Technology Research and Development Program (863 Program; Grant No. 2014AA020803), the National Science and Technology Major Projects (2013ZX09103003-016), Shanghai Science and Technology Development Fund (No.12140902300 and No.12410707400), Shanghai Health System Outstanding Young Talent Training Plan (No.XYQ2013118), Emerging cutting-edge technology joint research projects of Shanghai (No.SHDC12012106) and Tongji University outstanding youth programme (No. 1501219074). In addition, the present work was supported by the grants R01 CA72851, CA181572 and CA184792 from the National Cancer Institute, National Institutes of Health, and funds from the Baylor Research Institute.
Competing interests None declared.
Patient consent Obtained.
Ethics approval Institutional Review Boards (IRBs) of all participating institutions.
Provenance and peer review Not commissioned; externally peer reviewed.