

Abstract
OBJECTIVE To investigate apoptosis in human osteosarcoma MG63 cells induced by resveratrol and the molecular mechanism involved.
METHODS MG63 cells were treated with different concentrations of resveratrol and transmission electron microscopy was used to observe morphological changes occurring in apoptosis. The MTT method was used to determine the inhibitory rate and flow cytometry was used to assess apoptosis and to analyze the expression of the p21cip1/WAF1 and survivin proteins; the expression of p21cip1/WAF1 and survivin mRNAs was analyzed by the reverse transcriptase polymerase chain reaction (RT-PCR).
RESULTS After resveratrol treatment, the growth of the MG63 cells was significantly inhibited in a time- and dose-dependent fashion. By transmission electron microscopy, the cells displayed morphological changes characteristic of apoptosis, including formation of cytoplasmic vacuoles, chromatin condensation and margination. Flow cytometry showed that the growth of the cells was inhibited after resveratrol (10 mg/L and 20 mg/L) treatment. The inhibitory rates were (11.9 ± 0.63)% and (19.7 ± 0.88)% respectively. The quantity of treated cells in G0/G1 transition was increased, but the number in the S phase and G2/M transition was decreased. A subdiploid peak was observed. The expression of p21cip1/WAF1 was up-regulated while survivin was down-regulated.
CONCLUSION Resveratrol can inhibit growth and induce apoptosis of MG63 cells. Its molecular mechanism might be related to modulation of survivin and p21cip1/WAF1 expression.
keywords
Introduction
Osteosarcoma is the most common malignant bone tumor with its primary appearance during the second and third decades of life. Although an increased number of molecular studies have been initiated in recent years, no clear results or new therapeutic treatments have been devised [1]. Much effort has been focused on the discovery and development of new chemopreventive agents, especially agents targeted at events known to be involved in the process of carcinogenesis. Therefore, we have sought to identify novel agents that can effectively prevent carcinogenesis, but with minimal toxicity.
Naturally occurring dietary compounds such as resveratrol have gained considerable attention as cancer chemopreventive agents. Resveratrol, a polyphenolic phytoalexin found in grapes and other food products, was purified and shown to have cancer chemopreventive activity in assays representing the major stages of carcinogenesis. Results have suggested that resveratrol, a common constituent in the human diet, merited investigation as a potential cancer chemopreventive agent[2]. Some reports have shown that resveratrol is a potential cancer chemopreventive agent, as reports have shown that it can prevent tumor growth and metastasis of human lung carcinoma, prostate, pancreatic and liver cancers, and in breast cancer models[3-7].
However, the molecular mechanism underlying antitumorigenic or chemopreventive activities of this phytochemical remains largely unknown. This study was designed to explore the mechanism of resveratrol-induced apoptosis in MG63 cells. We have shown, for the first time to our knowledge, that resveratrol can inhibit the growth and induce apoptosis of human osteosarcoma MG63 cells.
Materials and Methods
Drugs and reagents
Resveratrol, propidium iodide (PI) and MTT were purchased from the Sigma Chemical Co (USA). A stock solution of resveratrol was made in dimethylsulfoxide (DMSO, Sigma, USA) at a concentration of 100 g/L and stored at -20 °C. Working dilutions were directly made in the tissue culture medium. The final concentration of DMSO was less than 0.1%. Cultures of control cells contained DMSO (0.1%) alone at an equivalent dilution to that in cultures containing resveratrol. Mouse anti-human p21cip1/WAF1 and survivin monoclonal antibodies and goat anti-mouse FITC-conjugated secondary antibodies were purchased from Santa Cruz Biotechnology (USA). Taq polymerase and AMV reverse transcriptase were purchased from promega (USA). TRIZOL reagent and other reagents for RT-PCR were obtained from Dingguo Biotechnology (China).
The cell line of MG63 human osteosarcoma was obtained from the China Center for Typical Culture Collection (Wuhan, China). The cells were maintained in modified Eagle medium (MEM, Gibco, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS), 100 U/ml penicillin and 100 μg/ml streptomycin, and incubated in a humidified atmosphere with 5% CO2 at 37°C. The media were changed every 3 d, and the cells were routinely sub-cultured using trypsin/ethylenedinitrilo-tetraacetic acid (EDTA) solution (Sigma) when they reached 90% confluence.
MTT assay
The cell viability was determined by the MTT assay. Briefly, the logarithmically growing MG63 cells were plated into a 96-well plate at a density of 1 × 104 cells/well. After 24 h, the supernatant was removed and 200 μl of different concentrations of the drugs were added to the cells (5, 10, 20, 30, and 40 mg/L) for 24 h, 48 h and 72 h, respectively. Control wells were treated with 0.1% DMSO alone. Then, 20 μl MTT (5 g/L) was added to each well followed by incubation for an additional 4 h. The supernatant was then removed and 150 μl DMSO added and the cells vibrated for 15 min. When the blue crystals were dissolved, the optical density (OD) was determined at 492 nm using a 96-well multiscanner autoreader (Anthos ht II, Austria). The inhibitory rate (IR) was calculated as follows: IR (%) = [1 - (OD of the experimental samples/OD of the control)] × 100%. The 50% inhibitory concentration (IC50) was calculated from the linear equation, which was deduced from a concentration versus survival rate regression curve (LOGIT method).
Morphologic analysis by transmission electron microscopy
MG63 cells treated with 20 mg/L resveratrol for 72 h were washed with PBS, centrifuged, and pre-fixed with 2.5% glutaraldehyde at 4°C overnight. The cells were then rinsed thoroughly in PBS and postfixed for 1 h with 1% osmium tetroxide, then dehydrated through a graded ethanol series, and embedded in Epon 812 (USA). The ultrastructure of the cells was analyzed in ultrathin sections under a transmission electron microscope (H-7500, Hitachi, Japan) after the sections were stained with uranyl acetate and lead citrate.
Flow cytometric analysis
MG63 cells were grown at 60%~70% confluence in 50 ml flasks, serum-starved for 24 h and then treated with 0, 10 and 20 mg/L resveratrol for 72 h, At the end of the treatment, adherent and floating cells were combined, centrifuged and washed twice with PBS. The cells were fixed with 2 ml of ice-cold 70% ethanol at 4°C for 2 h. After fixation, the cells were washed with PBS and resuspended in 0.5 ml propidium iodide (PI)/RNase A solution followed by incubation in the dark at room temperature for 15 min. The fluorescence emission of the stained cells was measured with a flow cytometer (EPICS-XL IV, Bechman Coulter, USA). Data were analyzed with Multipcycle software and a histogram of DNA content (x-axis, PI-fluorescence) versus counts (y-axis) produced.
RT-PCR for p21cip1/WAF1 and survivin mRNA expression
After the MG63 cells were treated with 0, 10 and 20 mg/L resveratrol for 72 h, total cellular RNA was extracted with the TRIZOL reagent according to the manufacturer’s instructions and quantified by ultraviolet (UV) absorbance spectroscopy. The reverse transcriptase reaction (total volume 20 μl) was conducted using the following mixture containing 1 μl 10 mmol/L dNTP, 4 μl 3 U RNase inhibitor, 1 μl 50 ng/μL random primer, 2 μl AMV reverse transcriptase, 4 μl 5 × AMV buffer, 5 μl RNA template and 3 μl ddH2O. The reaction was conducted at 25°C for 10 min, 42°C for 1 h, heated to 95°C for 5 min to inactivate AMV reverse transcriptase and kept at 4°C for 5 min. The newly synthesized cDNA was amplified by PCR (MT PT220, MT Co, USA). Primers were as follows: survivin forward primer: 5'-TCT CAA GGA CCA CCG CAT CT-3'; reverse primer: 5’-GAC AGA AAG GAA AGC GCA ACC-3’ (229 bp); p21cip1/WAF1 forward primer: 5’-GTG AGC GAT GGA ACT TCG ACT-3’, reverse primer: 5’-CGA GGC ACA AGG GTA CAA GAC-3’ (229 bp); β-actin forward primer: 5’-GTG GGG CGC CCC AGG CAC CA-3’; reverse primer: 5’-CTC CTT AAT GTC ACG CAC GAT TTC-3’, the amplified PCR product was 540 bp. Gene primers were synthesized by the Beijing Oake Co. The 20 μl reaction mixture contained 4 μl of cDNA template, 25 mmol/L MgCl2 1.5 μl, 0.5 U of Taq polymerase 1 μl, and 0.7 μl 10 μmol/L of primer. PCR amplification was conducted under the following conditions: pre-denaturation 94°C for 5 min, then 30 cycles denaturation at 94 °C for 30 s, annealing at 56°C for 30 s, extension 72° C for 45 s, followed by 72°C for 5 min. PCR products were separated on 1.5% agarose gel (1 × TBE running buffer) stained with ethidium bromide and viewed under ultraviolet light. The fluorescence intensity of β-actin fragments served as a reference for the p21cip1/WAF1 and survivin fragments.
Analysis of p21cip1/WAF1and survivin proteins by flow cytometry
MG63 cells were treated with 0, 10 and 20 mg/L resveratrol for 72 h. At the end of the treatment, adherent and floating cells were combined, centrifuged and washed twice with PBS. The cells were fixed with 2 ml of ice cold 70% ethanol at 4°C for 2 h. The sample fluorescence staining was performed by the indirect immufluorescence labing method. The cells were washed once with PBS and incubated with mouse anti-human p21cip1/WAF1 and survivin monoclonal antibodies at room temperature for 30 min. The samples were then washed with PBS and incubated in the dark for 30 min with goat antimouse FITC-conjugated secondary antibodies, washed with PBS, centrifuged and resuspended with PBS. One sample was used as a negative control with PBS replacing the first/second antibody. One sample was used as isotype controls with only the second fluorescence antibody. A histogram plot of FITC-fluorescence intensity (in logarithmic fluorescence intensity) (x-axis) versus counts (y-axis) was produced by flow cytometry. A fluorescence index [FI, FI = (fluorescence intensity of sample protein expression - fluorescence intensity of isotype control)/fluorescence intensity of the normal control] was used to describe p21cip1/WAF1 and survivin protein expression.
Statistical analysis
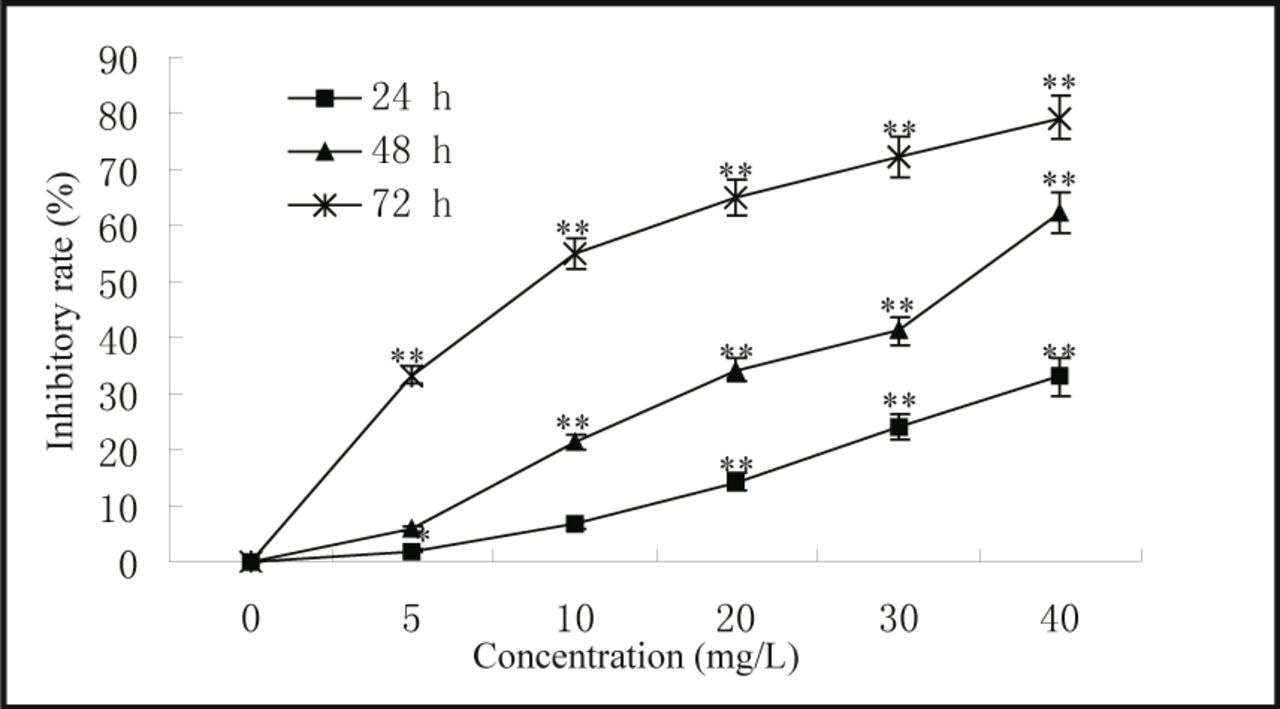
The experimental results were expressed as 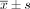 . Statistical analysis was conducted by one-way analysis of variance (ANOVA), and the differences between two groups were determined by the t-test using SPSS 12.0 statistical software. P < 0.05 was considered to be statistically significant, and P < 0.01 highly statistically significant.
. Statistical analysis was conducted by one-way analysis of variance (ANOVA), and the differences between two groups were determined by the t-test using SPSS 12.0 statistical software. P < 0.05 was considered to be statistically significant, and P < 0.01 highly statistically significant.
Results
Morphological changes of MG63 induced by resveratrol
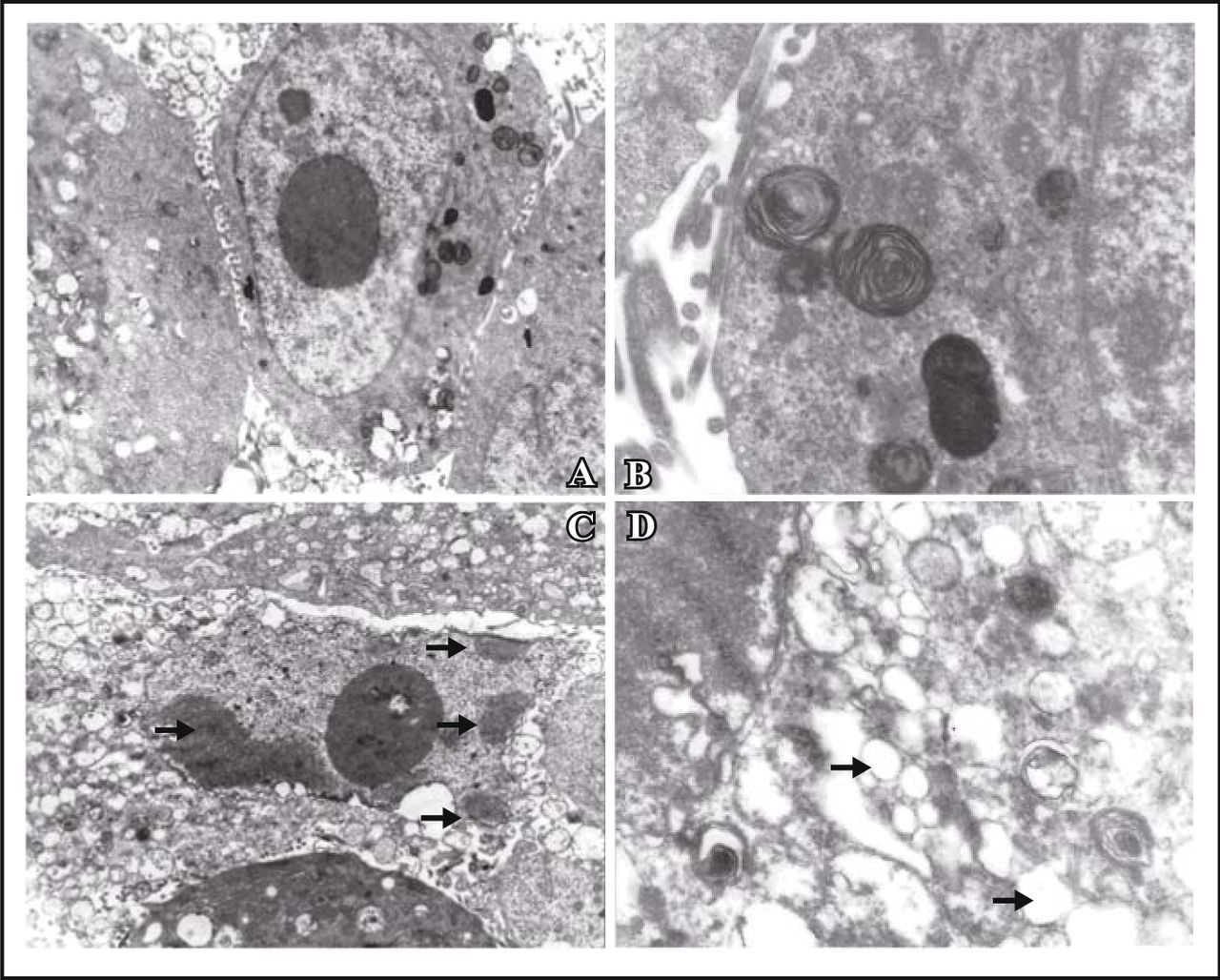
Under the transmission electron microscope, the control cells were big and round, with normal organelles, intact nuclear membrane, obvious nucleoli and low density in nuclear chromatin. There were rich microvilli on the surface (Fig.1.A&B). However, the cells treated with resveratrol exhibited the characteristics of apoptosis. The nuclear chromatin became condensed, marginated and segregated and the cytoplasm was vacuolated. The membrane of the cell, nucleus and cell organelles were all complete. Typical apoptotic bodies were not found (Fig.1.C&D).
MG63 cells showed apoptotic morphological changes of the chromatin such as being condensed and marginated (as showed by arrows in C ); There were lots of vacuolus in the cytoplasm (as shown by arrows in D). A, control cells × 4,000; B, control cells × 15,000; C, treated with resveratrol × 4,000; D, treated with resveratrol × 15,000.
Effect of resveratrol on growth inhibition in MG63 cell
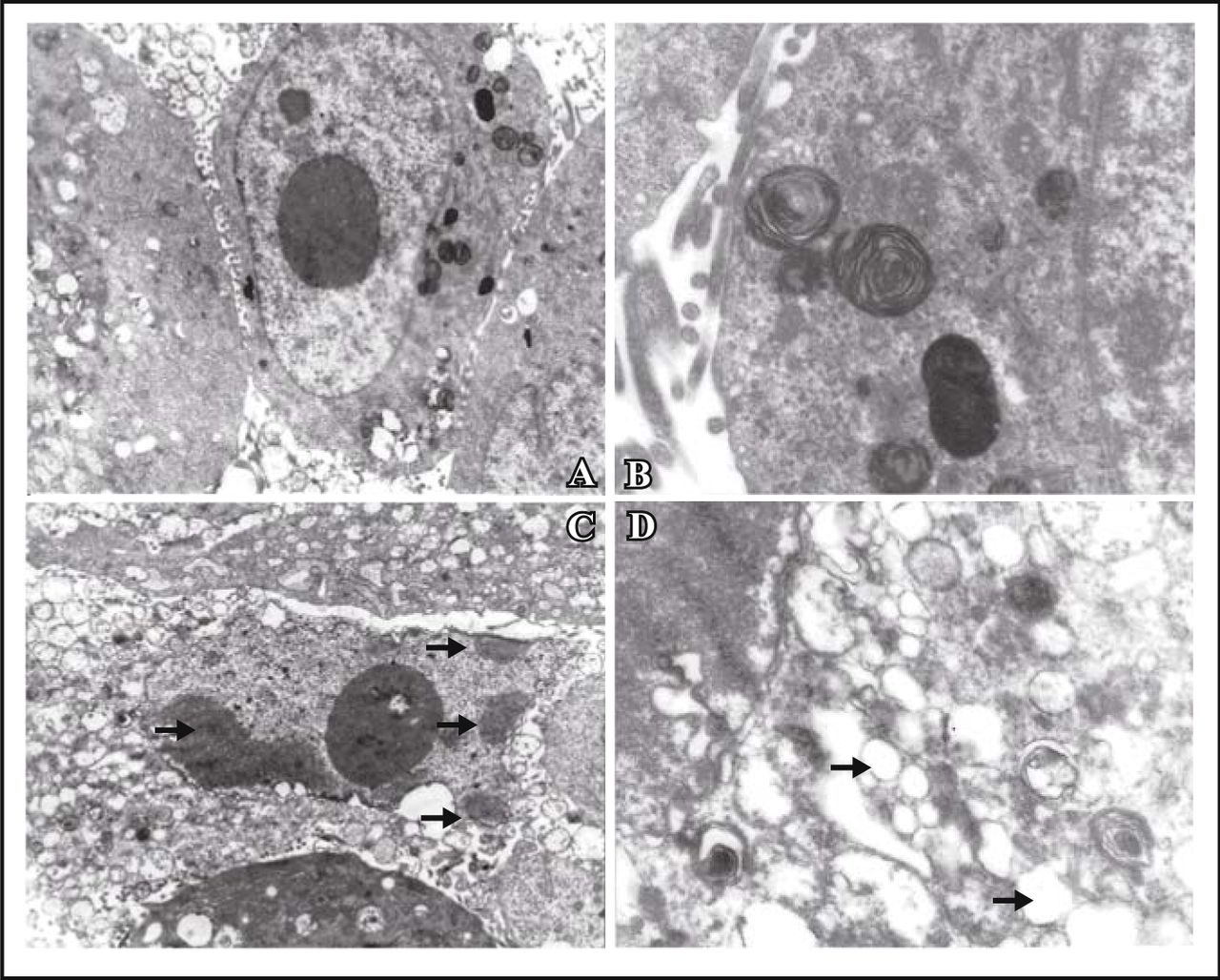
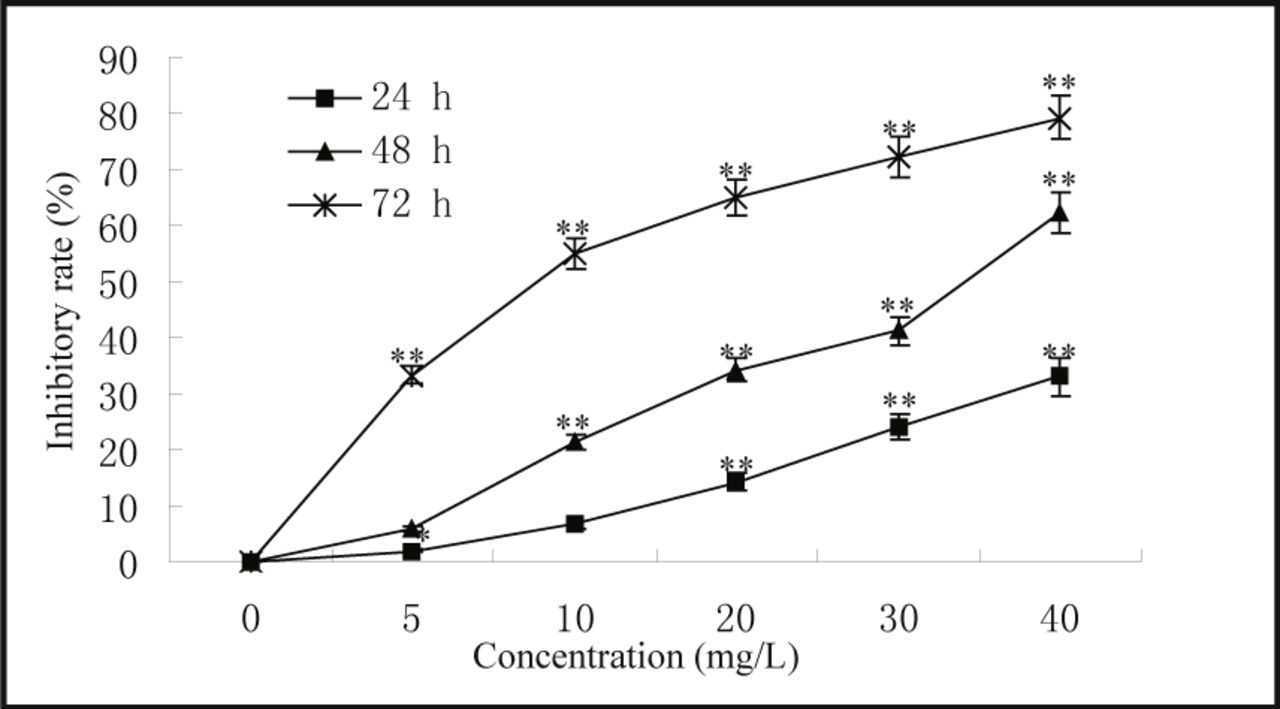
MG63 cells were treated with resveratrol at various concentrations for 24, 48, and 72 h, and the cell viability was determined as described by the MTT assay. The inhibitory effects of resveratrol were concentration and time-dependent. The biggest IR of resveratrol was 79.2% at the concentration of 40 mg/L at 72 h (Fig.2). It was found that the inhibitory effect was weak when cells were treated for 24 h. The IC50 at 48 h and 72 h were 31.4 mg/L and 9.69 mg/L respectively (Fig.2).
 ; n = 3; *, P < 0.05, **, P < 0.01; compared with control group.
; n = 3; *, P < 0.05, **, P < 0.01; compared with control group.
Effect of resveratrol on cell cycle parameters and apoptosis
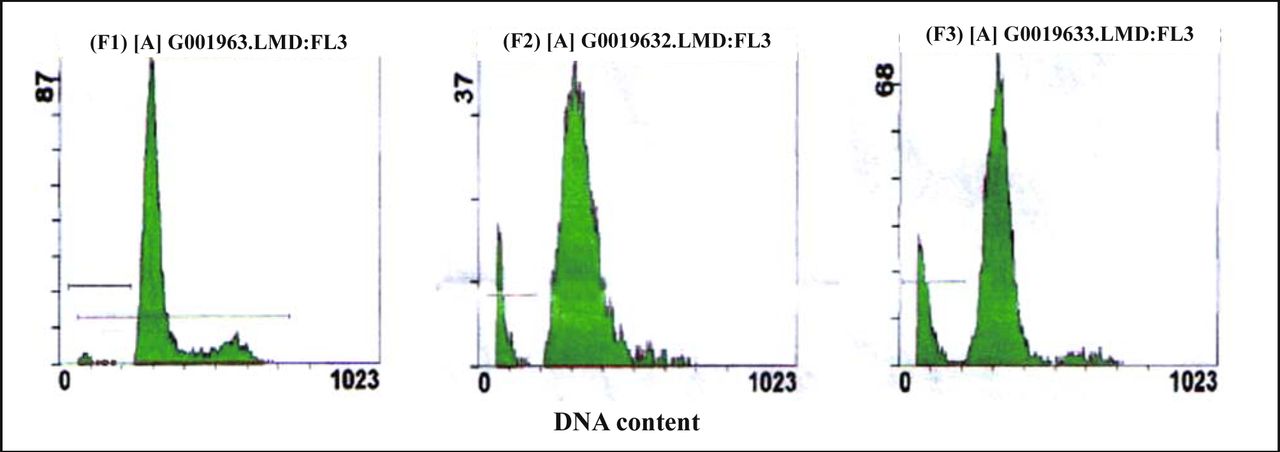
The analysis of cellular DNA content by FCM showed that there was a sub-G0/G1 peak in the graph of the drug-treated groups. That was a typical apoptotic peak, which did not show in the graph of control groups (Fig.3). After the MG63 cells were exposed to resveratrol (10 or 20 mg/L) for 72 h, the apoptotic rates were (11.90 ± 0.63)% and (19.85 ± 0.68)% respectively, which were significantly higher than the control (4.85 ± 0.21)% (P < 0.01) (Table 1,Fig.3). Most of the MG63 cells were blocked at the G0~G1 transition. Compared with the control group, cells at the G0~G1 transition of 20 mg/L treatment group increased by 13.3%, and cells in the S phase decreased by 10.1% (Table 1).
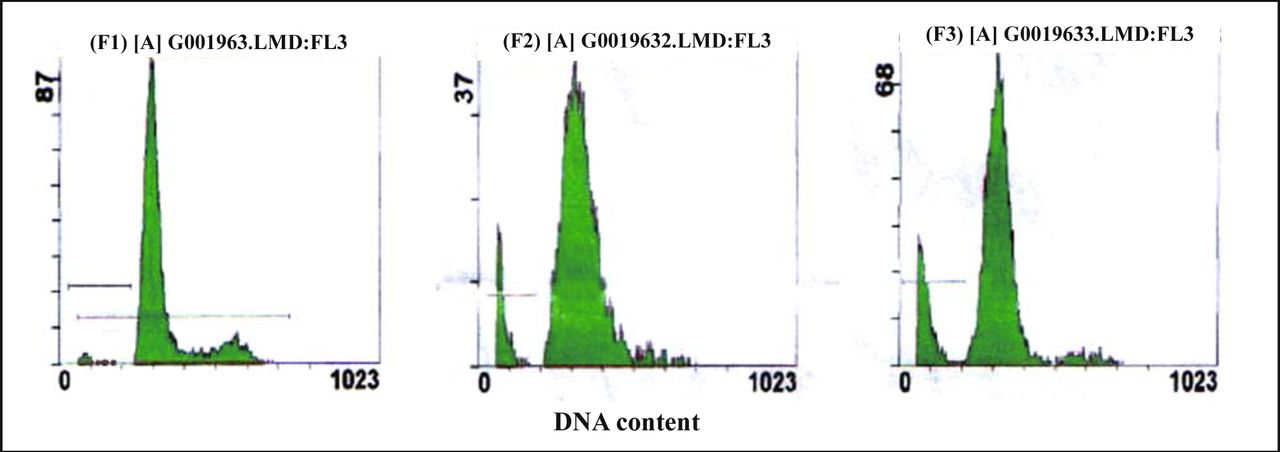
A, control cells; B&C, cells treated with resveratrol (10 or 20 mg/L) for 72 h, respectively. A sub-diploid peak was found before the G0/G1trasition, which was a typical apoptotic cell peak.
Effect of resveratrol on cell cycle parameters and apoptosis.
Effect of resveratrol on p21cip1/WAF1 and survivin mRNA expression in MG63 cells
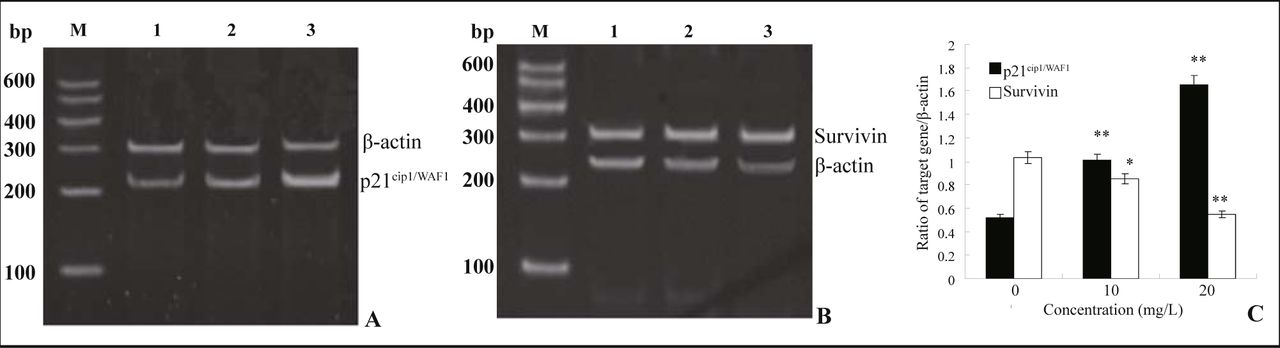
The MG63 cells were treated with resveratrol (10 or 20 mg/L) for 72 h, the p21cip1/WAF1 and survivin mRNAs were assessed by RT-PCR while β-actin was used as an internal control. In comparison with the control, the p21cip1/WAF1 mRNA expression level was dramatically increased (P < 0.01) (Fig.4. A&C). Control cells showed a basic expression of survivin and resveratrol treatmeat caused down-regulation of survivin mRNA expression (P < 0.05) (Fig.4. B&C).
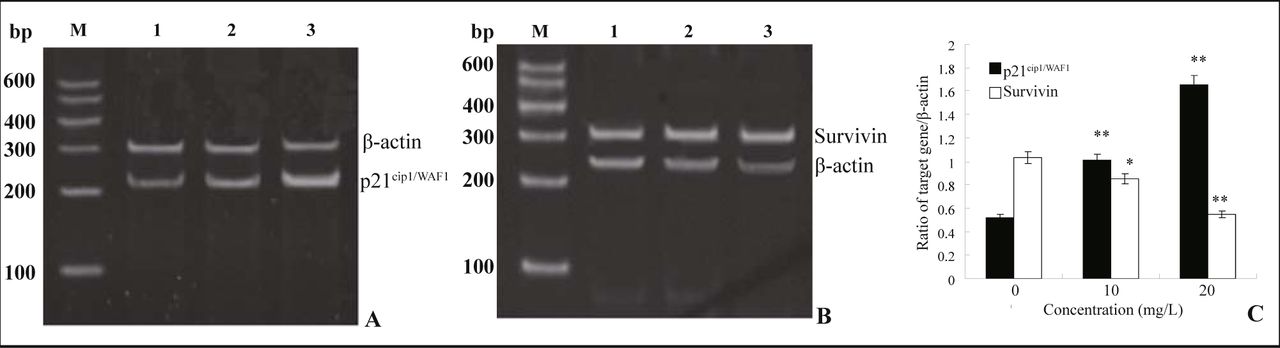
A and B show the mRNA expression of p21cip1/WAF1 and survivin respectively. Lane M, DNA marker; Lane 1, control cells; Lane 2, 10 mg/L resveratrol; Lane 3, 20 mg/L resveratrol. C shows the average of semi-quantitative amounts of p21cip1/WAF1 and survivin mRNA respectively. mRNA expression is presented as a ratio to internal standard. x ± s, n = 3, *P < 0.05, **P < 0.01, compared with control cells.
Expressions of p21cip1/WAF1 and survivin proteins
After MG63 cells were exposed to resveratrol (10 or 20 mg/L) for 72 h, FCM analysis showed that the expression of p21cip1/WAF1 was up-regulated while survivin was down-regulated (Table 2).
Effects of resveratrol on the expression of p21cip1/WAF1 and survivin.
Discussion
Osteosarcoma is the most common malignant bone tumor with approximately 1,000 new cases each year in the United States[1]. Due to neoadjuvant chemotherapy, the survival rate has increased to 60%~70% within the last 20 years. Regrettably, some groups of patients remain at high risk of eventual relapse[8].
Resveratrol, a trihydroxystilbene found in grapes and other plants, has been shown to be active in inhibiting multistage carcinogenesis. Our study was conducted to study the inhibitory effect of resveratrol on the proliferation of human osteosarcoma MG63 cells and to explore for possible mechanism. The results showed that resveratrol significantly inhibits cell growth in a time-and dose-dependent manner, with the biggest inhibitory effect reaching 79.01%. The drug-treated cells exhibited typical morphological apoptotic changes while the control cells showed normal morphology. The results of flow cytometry revealed that resveratrol induced apoptosis and caused arrest at the G0/G1 transition of the MG63 cells.
Cell cycle progression is controlled by various cyclin-dependent kinases (CDKs), whose activation is carefully regulated at multiple levels including the induction and degradation of cyclin protein, CDKs phosphorylation by cyclin-activating kinase, and the induction of CDK inhibitors (CDKIs)[9]. p21cip1/WAF1 was first cloned and characterized as an important effector that acts to inhibit CDK activity in p53-mediated cell cycle arrest in response to various agents[10]. The effects of resveratrol on the cell cycle have been demonstrated to be dependent on p21cip1/WAF1[11-13]. Our research aslo found resveratrol induced G0/G1 transition arrest and up-regulation of p21cip1/WAF1 expression in MG63 cells. These data indicate that up-regulation of p21cip1/WAF1 might play some role in the G0/G1 transition arrest induced by resveratrol.
We further tested the effect of resveratrol on the antiapoptosis gene, survivin. Survivin is a novel member of the apoptosis inhibitor gene family, which was identified by hybridization screening of human genome libraries with cDNA of effector cell protease receptor-1 (EPR-1) by Ambrosini et al.[14] at Yale University in 1997. Interestingly, survivin is expressed during embryonal development, in most human tumor tissues, and in many transformed cell lines, but not in normal adult tissues[15]. Trieb et al.[16] studied the localization and distribution of survivin in high-grade osteosarcomas by an indirect immunoperoxidase method. Results showed survivin expressioned in the cytoplasm or in the nucleus. Aziz MH et al.[17] evaluated the chemopreventive effects of resveratrol against ultraviolet-B (UVB) radiation-mediated skin tumorigenesis in the SKH-1 hairless mouse model. Results demonstrated that resveratrol imparted strong chemopreventive effects (relevant to human skin cancers) and that the chemopreventive effects of resveratrol might, at least in part, be mediated via modulations in survivin and other associated events. In our study, treatment with resveratrol resulted in reduction of survivin mRNA expression and survivin protein expression. In a more general application, the dietary compound resveratrol might serve as a novel therapeutic agent to target survivin expression in cancers through p21cip1/WAF1 mediated cell cycle arrest.
Apoptosis is a complex process involving multi-gene interactions, and unknown steps still remain in the whole pathway. Calcium is an intracellular second messenger, which acts in a spectrum of cellular processes ranging from fertilization to apoptosis. Many researches have indicated that an increase in intracellular calcium concentration could induce cell apoptosis[18,19], while some researches have shown that a reduction in cell calcium levels can induces apoptosis[20,21]. Our previous studies have investigated the effects of resveratrol on intracellular calcium concentration in rat ventricular myocytes. Laser scanning confocal microscopy and a whole cell patch-clamp recording technique show that resveratrol can reduce calcium influx[22,23]. The changes in the intracellular calcium concentration in MG63 cells treated with resveratrol need further investigation.
In conclusion, this study presents the first evidence that resveratrol can inhibit cell growth and induce apoptosis in MG63 cells. The mechanism might be related to increased expression of p21cip1/WAF1 and decreased expression of survivin.
- Received October 31, 2007.
- Accepted July 1, 2008.
- Copyright © 2008 by Tianjin Medical University Cancer Institute & Hospital and Springer
References
In this issue






{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.